



- Download PDF
- |
- Download Citation
- |
- Email a Colleague
- |
- Share:
-
- Tweet
-
Journal of Hematology and Therapeutics
Volume 1, Issue 2, October 2016, Pages 7–15
Original researchOpen Access
In vitro chemosensitivity study of human leukemic cells in a three-dimensional bone marrow culture system
-
Nobutada Fukino1,
Tomonori Harada1,*
 ,
Isao Tsuboi1,
Shinsuke Fukui2,
Masahiro Yasuda2and
Shin Aizawa1
,
Isao Tsuboi1,
Shinsuke Fukui2,
Masahiro Yasuda2and
Shin Aizawa1
*Corresponding author: Tomonori HARADA, M.D., Department of Functional Morphology, Nihon University School of Medicine, 30-1 Oyaguchikamicho, Itabashi-ku, Tokyo 173-8610, Japan. Tel.: +81-3-3972-8111 ext. 2222; Fax: +81-3-3973-8832; E-mail: harada.tomonori@nihon-u.ac.jp
Received 25 July 2016 Revised 15 September 2016 Accepted 20 September 2016 Published 27 September 2016
DOI: http://dx.doi.org/10.14312/2397-8694.2016-3
Copyright: © 2016 Tomonori HARADA, et al. Published by NobleResearch Publishers. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
AbstractTop
In the bone marrow, hematopoietic stem cell proliferation and differentiation are regulated by the hematopoietic microenvironment. This is reflected by the ability of the bone marrow microenvironment to alter the growth of leukemic cells, and protect leukemic cells from anti-cancer agents. Previously, we established a three-dimensional (3D) bone marrow culture system that maintained normal hematopoiesis, including prolongation of hematopoietic stem cell proliferation and differentiation. In the present study, we analyzed the effects of the anti-cancer drug, cytarabine, on the human leukemic cell line (K562) co-cultured with stromal cells in the 3D system. Comparisons were made with K562 cells treated with cytarabine in suspension or grown on a two-dimensional stromal cell monolayer (2D). We demonstrated that K562 cells cultured in the 3D system were more resistant to cytarabine treatment compared with cells grown in 2D or in suspension. Furthermore, there was a significant increase in the number of K562 cells in G0/G1 phase in 3D culture compared with cells grown in 2D or suspension cultures. These findings suggest that the differential response to cytarabine treatment in 3D culture may be related to the cell cycle period, which was modulated by stromal cells in the 3D microenvironment. Thus, the 3D culture system may be a valuable new tool for investigating leukemic cell-stromal cell interactions and the leukemic cell response to anti-cancer agents in vitro.
Keywords: 3D culture; bone marrow stromal cell; cell cycle; leukemia; cytarabine
IntroductionTop
Hematopoietic stem cells (HSCs) can self-renew and produce progenitors that are committed to differentiate in to a wide range of blood cell types, including erythrocytes, leukocytes, lymphocytes, and platelets. Earlier studies demonstrated that the proliferation and differentiation of HSCs are regulated by the bone marrow microenvironment [1-2]. Stromal cells, which are distinct from hematopoietic cells, are an essential component of this microenvironment, and are necessary for the long-term maintenance of HSCs in vitro [4, 5]. Several reports have shown that stromal cells regulate the proliferation and differentiation of HSCs through the production of diffusible hematopoietic regulatory factors and extracellular matrix, as well as through physical cell-cell interactions involving adhesion molecules and gap junction-mediated cell communication [1, 6-12]. Furthermore, the physiological function of stromal cells requires an anatomically appropriate three-dimensional (3D) orientation to determine HSC fate, which is linked to a normal and/or dysfunctional hematopoietic system [10, 13-16].
Stromal cells within the bone marrow microenvironment influence the proliferation of leukemic cells, as well as the normal hematopoietic cells from which they were derived. In addition, stromal cells likely protect leukemic cells from the effects of cytotoxic anti-cancer agents [17, 18]. In fact, current in vitro preclinical anti-cancer drug testing does not always predict in vivo responses [19]. Furthermore, even among patients who achieve complete remission with initial chemotherapy, the vast majority of patients with leukemia relapse because of residual disease in the bone marrow [20, 21]. However, the ability of stromal cells to support the proliferation (and differentiation) of HSCs or leukemic cells and to maintain their self-renewal potential has generally been investigated in long-term, two-dimensional (2D) bone marrow culture systems, which are structurally fundamentally different from the native hematopoietic environment. Hematopoiesis in vivo takes place in the bone marrow, where hematopoietic or leukemic cell proliferation is intimately linked with the 3D hematopoietic microenvironment, the so-called hematopoietic niche, which is composed of stromal cells. HSCs or leukemic cells are found in spaces within the bone marrow, and they rely on cell-cell interactions and the local milieu to determine their immediate fate [22].
We recently developed novel polymer particles with grafted epoxy polymer chains to support cell immobilization in a new 3D cell cultivation system [23-25]. The base polymer particles were synthesized by suspension polymerization of acrylic monomer and 2,2’-azobis[N-(2-propenyl)-2-methylpropionamide] (APMPA), and the epoxy polymer chain was extended from the particle surface by graft polymerization. The advantage of these particles is that the particle size, length and number of chains and the composition of the base polymer can be easily manipulated. The unique fibroblast cell line, MS-5, which was established from irradiated mouse bone marrow stromal cells and supports both murine and human HSC proliferation and differentiation in vitro [26-29], was used as a source of stromal cells in our 3D culture. The 3D cell bridges formed by MS-5 cells in the 3D cell cultivation system likely overcome the peeling problem associated with previous 3D and 2D systems, allowing long-term maintenance of HSCs in vitro [23]. Most HSCs in the 3D culture adhered (homed) to stromal cells, remained in the resting state, established an equilibrium between proliferation and differentiation, and similar to what is seen in hematopoietic tissue in vivo.
In this study, we designed our experiments to address the regulation of leukemic cell proliferation in the hematopoietic niche in 3D culture. As part of our approach, we examined the kinetics of leukemic cell growth in the presence of the anti-cancer drug, cytarabine (Ara-C), in our 3D culture system compared with 2D culture models. The combination therapy of cytarabine and anthracycline has been the standard and first line anti-leukemic regimen for more than 30 years [30]. Cytarabine is a nucleoside analog that is activated by phosphorylation to form cytosine arabinoside triphosphate (Ara-CTP), which competes with deoxycytidine triphosphate (dCTP) for incorporation into DNA. Thus, the anti-cancer effect of cytarabine depends on the cell cycle of target cells. The K562 human leukemic cell line was co-cultivated with MS-5 stromal cells, and the influence of cytarabine on the growth and cell cycle of K562 cells were assessed.
Materials and methodsTop
Materials
2,2’-azobis (isobutyronitrile), APMPA, dipotassium hydrogen phosphate, glutaraldehyde aqueous solution (25% w/v), glycidyl methacrylate (GMA), hydrochloric acid, methacrylic acid (MA), methanol, methyl methacrylate, paraformaldehyde aqueous solution (36% w/v), saccharose, sodium dihydrogen phosphate, and trypan blue were purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). Cresol red, poly (vinylpyrolidone) K-90, sodium hydroxide, and toluene were purchased from Nacalai Tesque (Kyoto, Japan). Pentaerythritol triacrylate was purchased from Sigma-Aldrich Co. (St. Louis, MO, USA).
Cytarabine was purchased from Sigma-Aldrich Co., dissolved in pyrogen-free saline to produce a concentration of 1 mg/mL, then diluted with Iscove’s modified Dulbecco’s medium (IMDM) for use. A previous study showed that the half-maximal effective concentration (EC50) of cytarabine for K562 cells was 0.064 µM (15 ng/mL) [31]. In the present study, cytarabine was used at concentrations ranging from 1 ng/mL (0.004 µM) to 10 µg/mL (40.1 µM).
Preparation of cell support with grafted polymer chains
Polymer particles with grafted epoxy polymer chains were prepared as described previously [24]. For graft polymerization, GMA and MA were used at a ratio of 4:1 (w/w), respectively, and the resulting particles were washed in a funnel with a 40 × volume of distilled water and methanol. These polymer particles are subsequently referred to as G-02 polymer particles.
The distribution of particle diameters was measured using the laser particle diameter analyzer Microtrac FRA (Microtrac Inc., Montgomery, PA, USA), and 150-200 µm particles were used in this study. The amount of epoxy group in the polymer particles was measured using the hydrochloric acid-dioxane method, and particles with more than 1.03 × 10−3 µmol/g-particle of epoxy group were selected [23].
Cultivation of MS-5 stromal cells and K562 cells
The murine stromal cell line, MS-5, was maintained in 7 mL IMDM supplemented with 10% fetal calf serum (FCS; Hyclone, Logan, UT, USA), penicillin (50 U/mL; Gibco BRL, Grand Island, NY, USA), and streptomycin (100 µg/mL; Gibco BRL) in 25 cm2 flasks (Falcon 3013; Corning, One Riverfront Plaza, NY, USA). The cells were cultured in a humidified incubator at 37°C with 5% CO2, sub-cultured at a split ratio of 1:4 every 7 days using 0.25% trypsin plus 0.02% EDTA in phosphate-buffered saline (PBS), and maintained as described previously [23].
Human leukemic K562 cells [32] were maintained in 7 mL IMDM supplemented with 10% FCS, penicillin (50 U/mL), and streptomycin (100 µg/mL) in 25 cm2 flasks. The cells were cultured in a humidified incubator at 37°C and 5% CO2, and sub-cultured at a split ratio of 1:10 every 5 days. For all experiments, the cells were used in their logarithmic growth phase.
3D co-culture system
MS-5 cells (5-10 × 105) were added to 5 mL IMDM supplemented with 10% FCS in the presence of 1–5 × 104 G-02 polymer particles in a 14 mL round-bottomed polypropylene tube (Falcon 2006; Corning). The mixture was incubated in a humidified incubator at 37°C and 5% CO2 for 24 h, then transferred into 35 mm plastic dishes (Falcon 3046; Corning), and the incubation was continued. Once cells were immobilized on the surface of the particles, they remained attached, proliferated and formed bridges between the particles. The supernatant was replaced with fresh growth medium every 7 days. After 2-3 weeks, when the MS-5 cells had grown and formed a 3D layer in the culture dish, K562 cells (5 × 104/mL) suspended in 5 mL growth medium were inoculated onto the MS-5 cells. Control cultures included MS-5 cells grown in dishes without G-02 particles, and K562 cells inoculated onto MS-5 cells in traditional 2D cultures.
Treatment with cytarabine
Three different experiments were designed. In the first, various concentrations of cytarabine were added to the culture at the same time as K562 cell inoculation, and the cells were left to grow undisturbed for the duration of the experiment. In the second, cytarabine was added to the culture at the same time as K562 inoculation and the cells were harvested (including some K562 cells adhered to stroma) 5 days later, washed once with IMDM to remove cytarabine, and then re-inoculated into the cytarabine free culture. Residual K562 cells in the supernatant or adherent layer after 5 days cytarabine treatment were able to grow in cytarabine free culture conditions. In the third, K562 cells were inoculated into the culture 2 days before cytarabine treatment. Pre-inoculated K562 cells adhered to the MS-5 cells, and the effect of cytarabine on the adhered cells were investigated.
Cultured K562 cells were harvested at various intervals for analysis. The supernatant (including floating cells) and cells in the adherent layer were collected separately. Adherent K562 cells were easily dislodged from the MS-5 cells in both 2D and 3D culture without trypsin treatment, and were harvested by repeated pipetting. The harvested cells from the supernatant and adherent layers were counted using a hemocytometer, and viable cells were distinguished by trypan blue dye exclusion. An aliquot of cells was assayed for CD45 and APO2.7 surface antigen expression, and the cell cycle was analyzed by flow cytometry.
Cell surface antigen analysis
Harvested cells were washed with PBS containing 2% FCS and passed through a 35 µm filter (Cell Strainer; Falcon 352235) to remove the polymer particles and aggregated cells (most of which were stromal MS-5 cells). Cells (2 × 105) were suspended in 0.5 mL of PBS containing 2% FCS and 0.02% NaN3 and incubated with one or both of the following monoclonal anti bodies (mAbs) for 30 min at 4°C: Phycoerythrin (PE)-conjugated mAb APO2.7 (clone 2.7 A6A3) for detection of the APO2.7 antigen expressed by apoptotic cells (Becton Dickinson, San Jose, CA, USA) [33, 34], and fluorescein isothiocynate (FITC)-conjugated anti-human CD45 (clone H130, Becton Dickinson) for the detection of hematopoietic cells. The cells were washed three times with PBS and assessed by flow cytometry on a Cytomix FC500 (Beckman Coulter, Brea, CA, USA).
Cell cycle analysis
Harvested cells were washed with PBS and passed through a 35 µm filter to remove the polymer particles and aggregated cells, and cell cycle analysis was performed using with Coulter DNA PREP Reagents Kit (Beckman Coulter) in accordance with the manufacturer’s instructions [35]. Briefly, 2 × 105 cells suspended in 0.1 mL of PBS were treated with 0.1 mL of DNA Prep LPR reagent, vortexed, and 2 mL of DNA PrepStain was immediately added. After further vortexing, the cells were incubated for 25 min at room temperature in a dark box, and the cell cycle was assessed by flow cytometry (Cytomix FC500, Beckman Coulter).
Statistical analysis
The results are expressed as the mean ± SD of triplicate experiments. Differences between the means were determined using two-way analysis of variance (ANOVA), and p ≤ 0.05 was considered significant.
ResultsTop
Sensitivity of K562 and MS-5 cells to cytarabine treatment
K562 cells in suspension culture (stromal cell free culture) grow logarithmically in the absence of cytarabine, and reach a growth plateau after 5 days of culture. Introduction of 10 ng/mL of cytarabine suppressed the growth of K562 cells significantly in suspension (stromal cell free) culture, consistent with the known EC50 of cytarabine for K562 cells (0.064 µM, 15 ng/mL) (Figure 1a) [31]. At concentrations of 100 ng/ml or greater, growth was effectively completely abrogated (Figure 1a). Thus, cytarabine at the concentrations of 1 µg/mL and 10 ng/mL were used for the following co-culture experiments.
Stromal MS-5 cells were seeded into 35 mm plastic dishes with or without G-02 particles, and high dose cytarabine (10 or 100 µg/mL) was introduced when MS-5 cells formed a 2D or 3D stromal layer on the bottom of the culture dish. After 5 days of cytarabine treatment, cells were collected using 0.25% trypsin/ 0.02% EDTA in PBS, and viable cell number was determined by trypan blue dye exclusion (Figure 1b). More than 90% of MS-5 cells collected from 2D or 3D culture were viable after treatment with 10 or 100 µg/mL of cytarabine. Thus, MS-5 cells were resistant to cytarabine treatment once the cells had formed a stromal layer.
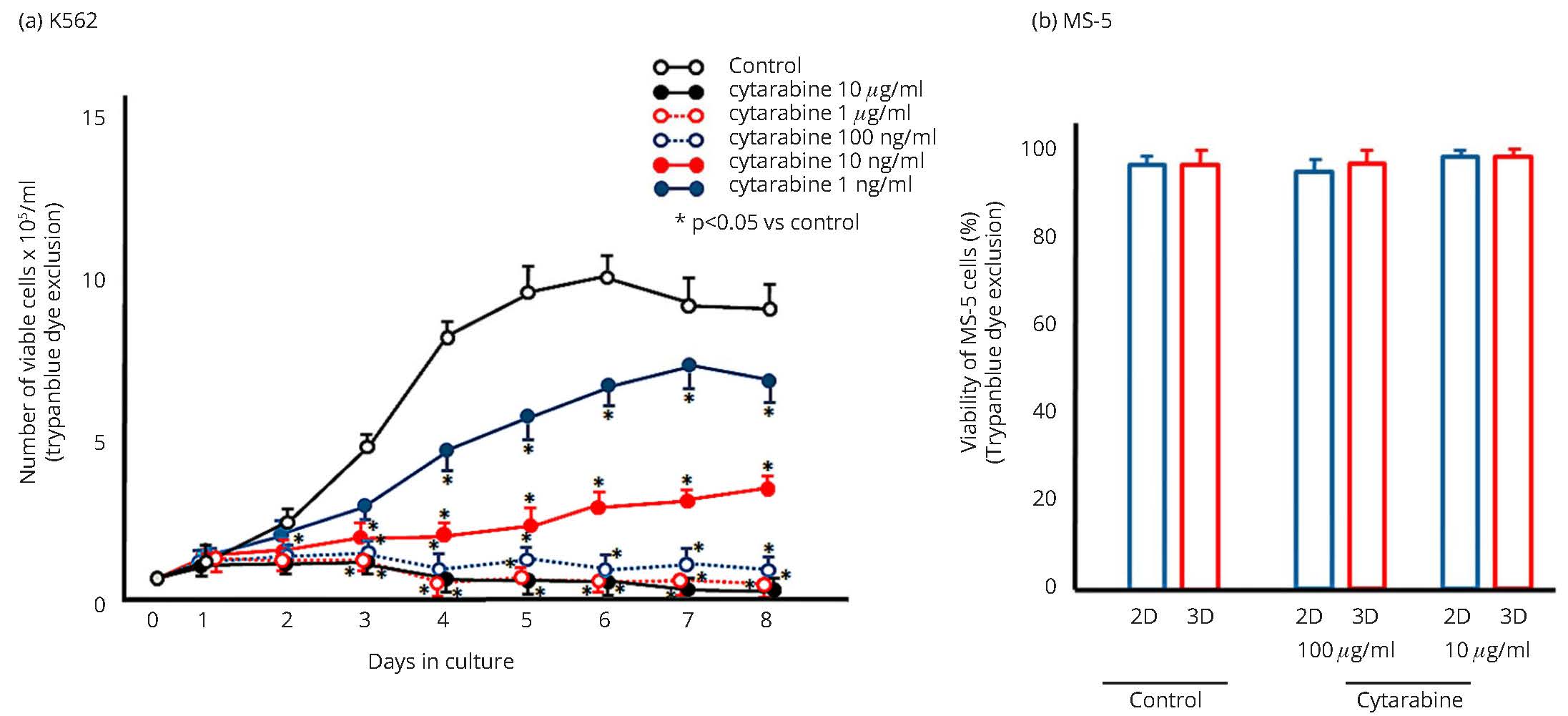
*Significant difference (p < 0.05) with respect to the cytarabine free control culture.
Influence of cytarabine on the growth of K562 cells in culture with stromal cells
K562 cells (5 × 104/mL) were inoculated into the 2D or 3D cultures. Figures 2a-c shows phase contrast images of K562 cells in stromal cell free, 2D, and 3D cultures, respectively. Inoculated K562 cells adhered to the surface of the flat MS-5 layer (2D culture, Figure 2b), and infiltrated the 3D structural MS-5 layer (3D culture, Figure 2c). After 5 days of cytarabine treatment (1 µg/mL) in stromal cell free or 2D culture, the shape of K562 cells become irregular with a slightly reduced cell-size (Figure 2d, e). In 3D culture, K562 cells remained adherent to the 3D structural MS-5 cells and maintained a rounded shape (Figure 2f).

The number of viable K562 cells was counted after cytarabine treatment. Firstly, 1 µg/mL or 10 ng/mL of cytarabine was added to the culture at the same time as K562 cell inoculation, and the viable cell number was counted after five days of cultivation (Figure 3a). In control cultures without cytarabine, the number of K562 cells in the 3D co-culture system was significantly lower than that without stromal cells, or in the 2D co-culture system. Although cytarabine reduced the number of K562 cells in all groups, the number of K562 cells in 3D co-culture was significantly higher than that in culture without stromal cells.
Next, 1 µg/mL of cytarabine was introduced into the culture at the same time as inoculation of K562 cells, and cells were collected after 5 days, washed, and re-cultured without cytarabine (Figure 3b). The surviving residual K562 cells began to proliferate in cytarabine free culture conditions. K562 cells in 3D culture were much more proliferative compared with the cells in stromal cell free or 2D culture, indicating that the K562 cells in 3D culture were more resistant to cytarabine treatment.
To examine the effect of cytarabine on adhered K562 cells, the drug was introduced 2 days after inoculation of K562 cells (Figure 3c), which allowed enough time for K562 cells to adhere to stromal MS-5 cells. The number of cells in 3D culture was significantly greater than that in stromal cell free or 2D culture. Furthermore, the number of K562 cells in 3D culture was higher than the number obtained when K562 cells and cytarabine were added simultaneously, indicating that once K562 cells adhered to the stroma the cytoreductive effect of cytarabine was diminished.
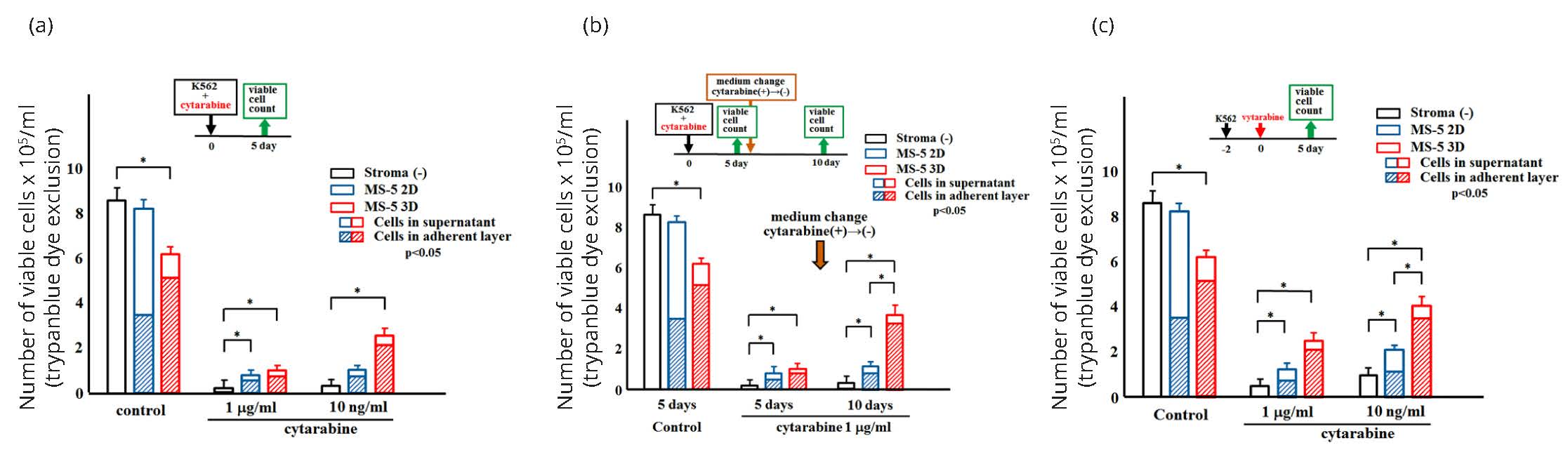
*Significant difference (p < 0.05).
Expression of the APO2.7 antigen on K562 cells after treatment with cytarabine
To address the mechanisms underlying the reductions in K562 cell numbers in response to cytarabine treatment, the expression of the APO2.7 apoptosis antigen was examined. An aliquot of cells was collected from each culture 5 days after treatment (using the same protocol as for Figure 3a) and double stained with anti-CD45 and APO2.7 antibodies (Figure 4). Human leukemic K562 cells were positive for CD45, and murine MS-5 cells were negative for CD45. Figure 4a shows the typical dot plot histogram of two-color cytometry determined by CD45 (FL-1) and Apo2.7 (FL-2). Figure 4b represents the typical flow cytometry histograms of K562 cells treated with cytarabine and stained for APO2.7, and the results are summarized in Figure 4c. Significantly higher numbers of APO2.7 positive cells were observed in culture without stromal cells, suggesting that apoptosis was induced under those conditions (Figure 4c). The lowest APO2.7 expression was observed on MS-5 adherent cells in 3D culture compared with floating cells and adherent cells in 2D culture (Figure 4d). Overall, these data demonstrate that in 3D culture MS-5 cells protect K562 cells from apoptosis induced by cytarabine treatment.
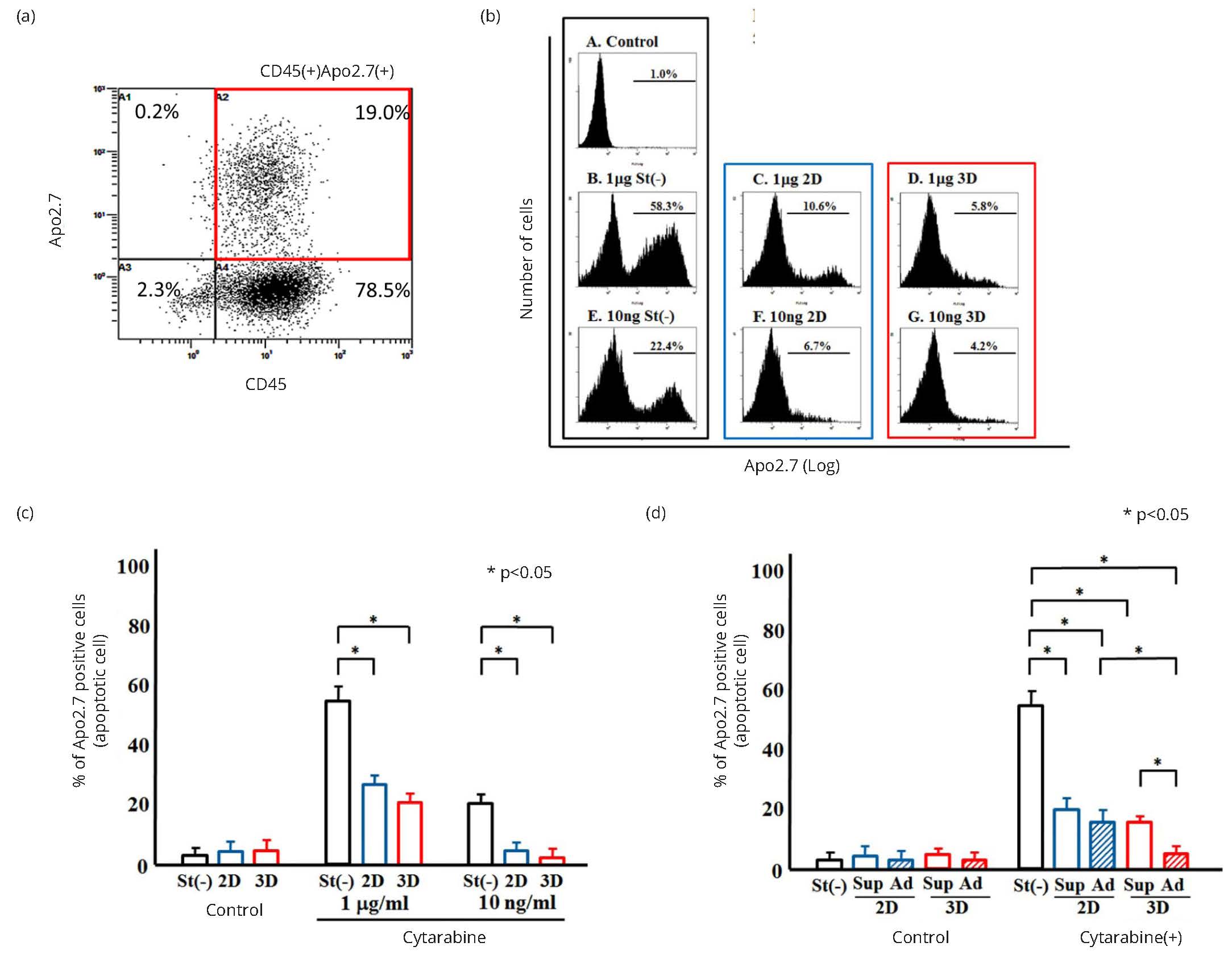
*Significant difference (p < 0.05)
Cell cycle analysis in cultured K562 cells
K562 cells were collected from stromal cell free, 2D, and 3D cultures without cytarabine treatment, and the cell cycle was analyzed by flow cytometry. Figure 5a shows the typical cell cycle pattern at day 1 of culture. The percentage of resting K562 cells (G0/G1) was 40.1 ± 2.8% in stromal cell free culture, 42.2 ± 3.2% in 2D culture and 61.4 ± 3.5% in 3D culture. The percentage of resting K562 cells in stroma free and 2D culture increased daily as K562 cell numbers increased, and no significant differences between cells collected from stroma free and 2D culture were observed at day 3 and 4 of culture (Figure 5b). On the other hand, the percentage of resting cells in 3D culture did not change over 4 days (Figure 5b), even though the cell number in culture increased (Figure 3). In addition, a higher percentage of adherent K562 cells were in the resting state compared with floating cells in 3D culture (Figure 6). K562 cells in 3D culture appeared to maintain a stable cell cycle, and more than 50% of cells were in resting phase at any one time.
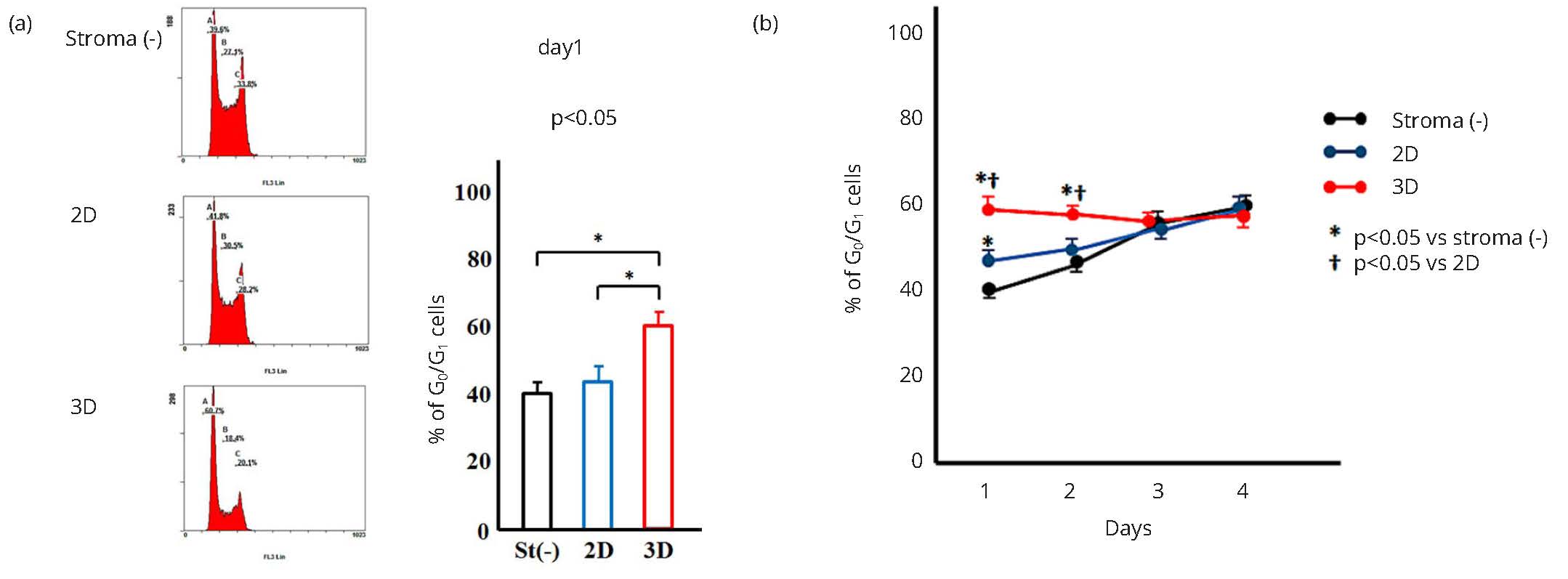
*Significant difference (p < 0.05)
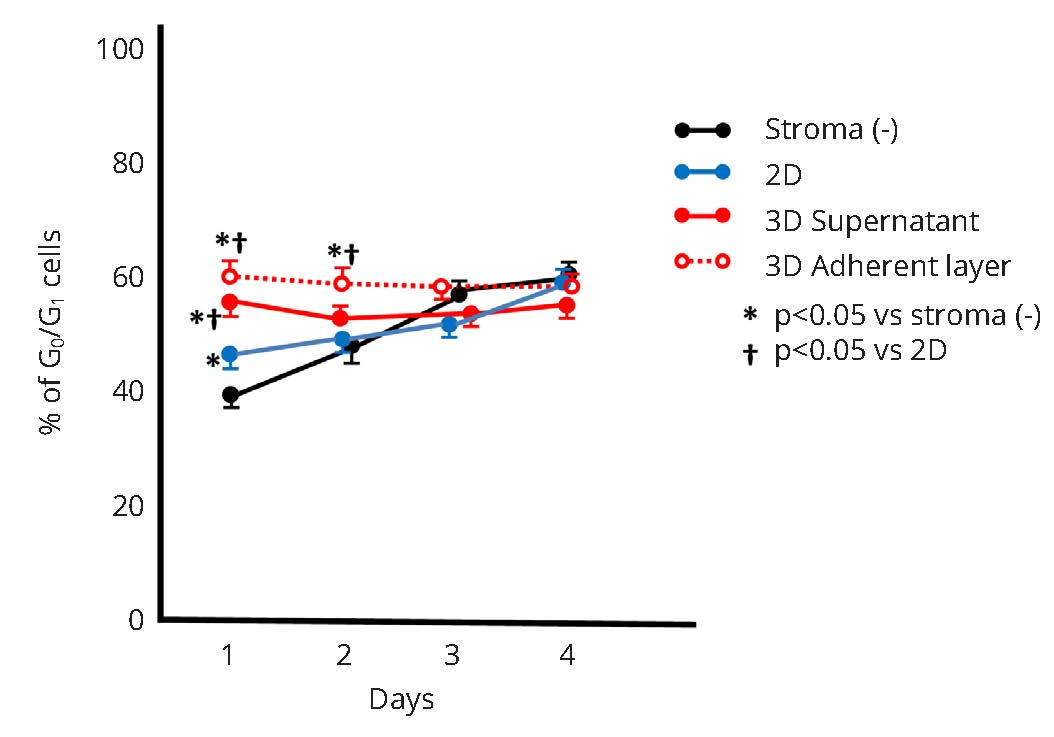
*Significant difference (p < 0.05).
DiscussionTop
In vivo, hematopoiesis takes place in the bone marrow, where hematopoietic cells are in intimate association with distinctive stromal cell elements. The results of several previous studies have suggested that the stromal cell microenvironment plays an important role in hematopoiesis [1-3], and Schofield proposed the concept of a “niche” as a specialized microenvironment housing HSCs [36]. A variety of in vitro co-culture experiments with stromal and hematopoietic cells demonstrated that both stromal cell function and niche anatomy regulate stem cell quiescence, self-renewal, and differentiation [6-9, 11, 12, 37, 38]. Abnormal clones originating from hematopoietic cells, including leukemic clones, also arise within the niche. Stromal cells in the niche may supervise and suppress the growth of abnormal clones to maintain normal hematopoiesis [34] by producing soluble factors and mediatory cell-to-cell interactions via adhesion molecules [39-41]. It is possible that if stromal cell regulatory mechanisms are compromised, hyper-proliferation of abnormal clones may occur. Thus, stromal cells regulate the proliferation of not only normal hematopoietic cells but also abnormal clones. However, the mechanisms underlying stromal cells regulatory functions remain unclear.
Previously, we established a novel 3D culture system using polymer particles with grafted epoxy polymer chains for cell immobilization [23, 24]. In our 3D culture system, MS-5 cells are used as stromal cells [23], and HSC proliferation and differentiation are maintained for significantly longer, recapitulating more closely the natural hematopoietic niche compared with traditional 2D culture systems. Interestingly, most HSCs were in the S phase in both the supernatant and adherent layer in 2D cultures at 4 weeks of co-culture, although more than 50% of those cells were outside S phase in the 3D cultures [25], indicating that the stromal cells in 3D culture control the proliferation and differentiation of hematopoietic cells by regulating the cell cycle. Similarly, the results from this study indicate that stromal cells also regulate the proliferation of leukemic cells in 3D culture, consistent with the importance of the hematopoietic niche in leukemogenesis.
Although our study did not address the mechanisms by which stromal cells regulate the K562 cell cycle and apoptosis, the data highlight the fact that the investigation of anti-cancer drugs in vitro is not straightforward. An unique 3D in vitro culture model was shown by Torisawa et al. [42]. They use a poly(dimethylsiloxane) (PDMS) device and histological examination showed the mimic cultured specimen to in vivo. This model showed the organ-level marrow protective effects to toxic agents, however, PDMS devise was necessary to implant to subcutaneously on the back of the mouse for 4-8 weeks prior to the culture, then cultured in special microfluidic system. In this regard, the practical advantages of our 3D culture system, including easy visual microscopic observation, and identification of human K562 leukemic cells and murine stromal cells by differential surface maker expression, make our system a particularly attractive new approach to address the complexities associated with the analysis of cancer cell chemosensitivity in vitro. In addition, our 3D co-culture system with stromal cells does not require the addition of any biological agents, such as hematopoietic growth factors to support hematopoietic and leukemic cell proliferation.
In relation to cytarabine sensitivity, our data accord with several previous studies that demonstrated that apoptotic changes induced by cytarabine in leukemic cell lines or primary cells were reduced by co-culture with stromal cells in 2D in vitro models. Lee et al. [43] and Macanas-Pirard et al. [44] demonstrated that soluble factor(s) produced by stromal cells protected leukemic cells from apoptosis induced by cytarabine treatment. In addition, Shishido et al. [45] and Kogoshi et al. [46] showed that stromal cells-mediated resistance to cytarabine treatment by leukemic cells through cell-to-cell interactions via adherent molecules. Recent 3D existing models showed that cell adhesion-mediated drug resistance protects the leukemic cells from apoptosis induce by cytotoxic agents. Damiano et al. [47] and Kuzelova et al. [48] reported that leukemic cells (e.g. K562 cells) adhered to fibronectin were found to be resistant to apoptosis induced by DNA damaging agents. Aljitawi et al. [49] reported that leukemic cells in 3D culture using Scaffold fibers, were resistant to drug-induced apoptosis compared to cells cultured in 2D, and chemo-resistant response of leukemic cells was related to the expression of N-cadherin in 3D culture system. In facts, Xishan et al. [50] also demonstrated that the influences of N-cadherin on K562 cell proliferation in vitro. All these models provide an opportunity to study leukemic cell responses to anti-cancer agents in 3D culture system. Importantly, our results revealed that although stromal MS-5 cells in 2D culture partially protected K562 cells from cytarabine induced apoptosis, greater protection was observed in our 3D system. This is the first report demonstrating that the capacity of stromal cells to mediate the resistance of leukemic cells to cytarabine is dependent on the culture system (2D and 3D). Why this is the case requires further investigation. However, we previously demonstrated that the expression of various genes involved in the regulation of hematopoietic cell proliferation and differentiation was “stabilized” in stromal cells in 3D culture compared to those in 2D culture, suggesting that 3D culture promotes the establishment of a “resting” niche in which most hematopoietic cells are outside S phase [25].
In the future, it will be necessary to clarify the mechanisms regulating hematopoietic or leukemic cell homing to the stromal niche, and the maintenance of the resting condition. Comparative studies of 2D and 3D cultures to investigate stromal cell production of various soluble factors, extracellular matrix [51] and adherent molecules may help to resolve these questions.
ConclusionTop
Stromal cells in our 3D culture system regulated leukemic (K562) cell proliferation by controlling the cell cycle. In 3D culture, most K562 cells were maintained in a stable resting state and the cytocidal activity of cytarabine was reduced compared with cells grown in 2D culture. Although further studies are required to analyze the mechanisms responsible for the cell cycle regulatory activity of MS-5 cells in 3D culture, the results indicate that our 3D culture system may be a valuable new tool for investigating the cell-to-cell interactions between stroma and leukemic cells in vitro.
Author contributions
All authors participated in the design, interpretation of the studies, analysis of the data, and review of the manuscript. NF, TH, IT, SF, MY and SA conducted the experiments, SF and MY made polymer particles, and NF, TH and SA wrote the manuscript.
Acknowledgment
This work was supported in part by JSPS KAKENHI Number 15K08161. We thank Ms. Sonoko Araki and Ms. Miyuki Yuda for their technical assistance.
Conflicts of interest
The authors have no conflicts of interest to declare.
ReferencesTop
[1]Greenberger JS. The hematopoietic microenvironment. Crit Rev Oncol Hematol. 1991; 11(1):65–84.Article Pubmed
[2]Mayani H, Guilbert LJ, Janowska-Wieczorek A. Biology of the hemopoietic microenvironment. Eur J Haematol. 1992; 49(5):225–233.Article Pubmed
[3]Wolf NS, Trentin JJ. Hemopoietic colony studies. V. Effect of hemopoietic organ stroma on differentiation of pluripotent stem cells. J Exp Med. 1968; 127(1):205–214.Article Pubmed
[4]Dexter TM, Allen TD, Lajtha LG. Conditions controlling the proliferation of haematopoietic stem cells in vitro. J Cell Physiol. 1977; 91(3):335–344.Article Pubmed
[5]Mori KJ, Fujitake H, Okubo H, Dexter TM, Ito Y. Quantitative development of adherent cell colonies in bone marrow cell culture in vitro. Exp Hematol. 1979; 7(4):171–176.Pubmed
[6]Aizawa S, Yaguchi M, Nakano M, Toyama K, Inokuchi S, Imai T, et al. Hemopoietic supportive function of human bone marrow stromal cell lines established by a recombinant SV40-adenovirus vector. Exp Hematol. 1994; 22(6):482–487.Pubmed
[7]Bodo M, Baroni T, Tabilio A. Haemopoietic and stromal stem cell regulation by extracellular matrix components and growth factors. J Stem Cells. 2009; 4(1):57–69.Article Pubmed
[8]Dormady SP, Bashayan O, Dougherty R, Zhang XM, Basch RS. Immortalized multipotential mesenchymal cells and the hematopoietic microenvironment. J Hematother Stem Cell Res. 2001; 10(1):125–140.Article Pubmed
[9]Durig J, Rosenthal C, Halfmeyer K, Wiemann M, Novotny J, et al. Intercellular communication between bone marrow stromal cells and CD34+ haematopoietic progenitor cells is mediated by connexin 43-type gap junctions. Br J Haematol. 2000; 111(2):416–425.Article Pubmed
[10]Garrett RW, Emerson SG. The role of parathyroid hormone and insulin-like growth factors in hematopoietic niches: physiology and pharmacology. Mol Cell Endocrinol. 2008; 288(1-2):6–10.Article Pubmed
[11]Heard J, Fichekson S, Varet B. Role of colony-stimulating activity in murine long-term bone marrow cultures: Evidence for its production and consumption by the adherent cells. Blood. 1982; 59(4):761–767.Article Pubmed
[12]Wagner W, Roderburg C, Wein F, Diehlmann A, Frankhauser M, et al. Molecular and secretory profiles of human mesenchymal stromal cells and their abilities to maintain primitive hematopoietic progenitors. Stem Cells. 2007; 25(10):2638–2647.Article Pubmed
[13]Can A. Haematopoietic stem cells niches: Interrelations between structure and function. Transfus Apher Sci. 2008; 38(3):261–268.Article Pubmed
[14]Himburg HA, Harris JR, Ito T, Daher P, Russell JL, et al. Pleiotrophin regulates the retention and self-renewal of hematopoietic stem cells in the bone marrow vascular niche. Cell Rep. 2012; 2(4):964–975.Article Pubmed
[15]Nagasawa T, Omatsu Y, Sugiyama T. Control of hematopoietic stem cells by the bone marrow stromal niche: The role of reticular cells. Trends Immunol. 2011; 32(7):315–320.Article Pubmed
[16]Renström J, Kröger M, Peschel C, Oostendorp RA. How the niche regulates hematopoietic stem cells. Chem Biol Interact. 2010; 184(1-2):7–15.Article Pubmed
[17]Li ZW, Dalton WS. Tumor microenvironment and drug resistance in hematologic malignancies. Blood Rev. 2006; 20(6):333–342.Article Pubmed
[18]Konopleva M, Konoplev S, Hu W, Zaritskey AY, Afanasiev BV, et al. Stromal cells prevent apoptosis of AML cells by up-regulation of anti-apoptotic proteins. Leukemia. 2002; 16(9):1713–1724.Article Pubmed
[19]Kunz-Schughart LA, Freyer JP, Hofstaedter F, Ebner R. The use of 3-D cultures for high-throughput screening: The multicellular spheroid model. J Biomol Screen. 2004; 9(4):273–85.Article Pubmed
[20]Tallman MS, Gilliland DG, Rowe JM. Drug therapy for acute myeloid leukemia. Blood. 2005; 106(4):1154–1163.Article Pubmed
[21]Ding L, Ley TJ, Larson DE, Miller CA, Koboldt DC, et al. Clonal evolution in relapsed acute myeloid leukaemia revealed by whole-genome sequencing. Nature. 2012; 481(7382):506–510.Article Pubmed
[22]Adams GB, Scadden DT. The hematopoietic stem cell in its place. Nat Immunol. 2006; 7(4):333–337.Article Pubmed
[23]Hirabayashi Y, Hatta Y, Takeuchi J, Tsuboi I, Harada T, et al. Novel three-dimensional long-term bone marrow culture system using polymer particles with grafted epoxy-polymer-chains supports the proliferation and differentiation of hematopoietic stem cells. Exp Biol Med. 2011; 236(11):1342–1350.Article Pubmed
[24]Yasuda M, Kunieda H, Ono K, Ogino H, Iwasaki T, et al. Adhesive cell cultivation on polymer particles having grafted epoxy polymer chain. Tissue Cell. 2011; 43(2):115–124.Article Pubmed
[25]Harada T, Hirabayashi Y, Hatta Y, Tsuboi I, Glomn WR, et al. Kinetics of hematopoietic stem cells and supportive activities of stromal cells in a three-dimensional bone marrow culture system. Growth Factors. 2015; 33(5-6):347–355.Article Pubmed
[26]Andersson LC, Nilsson K, Gahmberg CG. K562-a human erythroleukemic cell line. Int J Cancer. 1979; 23(2):143–147.Article Pubmed
[27]Hassan HT, Tsiritotis C, Maurer HR. Effects of recombinant human interferon-alpha, beta and gamma on the antiproliferative activity of cytarabine in K562 human myeloid leukemia clonogenic cells. Hematol Oncol. 1992; 10(2):117–124.Pubmed
[28]Zhang C, Ao Z, Seth A, Schlossman SF. A mitochondrial membrane protein defined by a nove monoclonal antibody is preferentially detected in apoptotic cells. J Immunol. 1996; 157(9):3980–3987.Article Pubmed
[29]Suzuki J, Fujita J, Taniguchi S, Sugimoto K, Mori KJ. Characterization of murine hemopoietic-supportive (MS-1 and MS-5) and non-supportive (MS-K) cell lines. Leukemia. 1992; 6(5):452–458.Article Pubmed
[30]Short NJ, Ravandi F. Acute Myeloid Leukemia: Past, present, and prospects for the future. Clin Lymphoma Myeloma Leuk. 2016; 16 Suppl:S25–29.Article Pubmed
[31]Hassan HT, Tsiriyotis C, Maurer HR. Effects of recombinant human interferon-alpha, beta and gamma on the antiproliferative activity of cytarabine in K562 human myeloid leukemia clonogenic cells. Hematol Oncol. 1992; 10(2):117–124.Pubmed
[32]Andersson LC, Nilsson K, Gahmberg CG. K562-a human erythroleukemic cell line. Int J Cancer. 1979; 23(2):143–147.Article Pubmed
[33]Zhang C, Ao Z, Seth A, Schlossman SF. A mitochondrial membrane protein defined by a nove monoclonal antibody is preferentially detected in apoptotic cells. J Immunol. 1996; 157(9):3980–3987.Article Pubmed
[34]Aizawa S, Hiramoto M, Hoshi H, Toyama K, Shima D, et al. Establishment of a stromal cell line from a MDS RA patient which induced an apoptotic change in hematopoietic and leukemic cells in vitro. Exp Hematol. 2000; 28(2):148–155.Article Pubmed
[35]Crissman HA, Steinkamp JA. Rapid, simultaneous measurement of DNA, protein, and cell volume in single cells from large mammalian cell populations. J Cell Biol. 1973; 59(3):766–771.Article Pubmed
[36]Schofield R. The relationship between the spleen colony-forming cell and the hematopoietic stem cell. Blood Cells. 1978; 4(1-2):7–25.Pubmed
[37]Paraguassu-Braga FH, Borojevic R, Bouzas LF, Barcinski MA, Bonomo A. Bone marrow stroma inhibits proliferation and apoptosis in leukemic cells through gap junction-mediated cell communication. Cell Death Differ. 2003; 10(9):1101–1108.Article Pubmed
[38]Wang X, Hisha H, Mizokami T, Cui W, Cui Y, et al. Mouse mesenchymal stem cells can support human hematopoiesis both in vitro and in vivo: the crucial role of neural cell adhesion molecule. Haematologica. 2010; 95(6):884–891.Article Pubmed
[39]Aikawa S, Hatta Y, Tanaka M, Kaneita Y, Yasukawa K, et al. Requirement of soluble factor(s) produced by bone marrow stromal cells on the growth of novel established human myeloma cell line. Int J Oncol. 2003; 22(3):631–637.Article Pubmed
[40]Chauhan D, Uchiyama H, Akbarali Y, Urashima M, Yamamoto K, et al. Multiple myeloma cell adhesion-induced interleukin-6 expression in bone marrow stromal cells involves activation of NF-kappa B. Blood. 1996; 87(3):1104–1112.Article Pubmed
[41]Kim I, Uchiyama H, Chauhan D, Anderson KC. Cell surface expression and functional significance of adhesion molecules on human myeloma-derived cell lines. Br J Haematol. 1994; 87(3):483–493.Article Pubmed
[42]Torisawa YS, Spina CS, Mammoto T, Mammoto A, Weaver JC, et al. Bone marrow-on-a-chip replicates hematopoietic niche physiology in vitro. Nat Methods. 2014; 11:663–669.Article Pubmed
[43]Lee YC, Chiou TJ, Tzeng WF, Chu ST. Macrophage inflammatory protein-3alpha influences growth of K562 leukemia cells in co-culture with anticancer drug-pretreated HS-5 stromal cells. Toxicology. 2008; 249(2-3):116–122.Article Pubmed
[44]Macanas-Pirard P, Leisewitz A, Broekhuizen R, Cautivo K, Barriga FM, et al. Bone marrow stromal cells modulate mouse ENT1 activity and protect leukemia cells from cytarabine induced apoptosis. PLoS One. 2012; 7 (5):e37203.Article Pubmed
[45]Shishido S, Bonig H, Kim YM. Role of integrin alpha4 in drug resistance of leukemia. Front Oncol. 2014; 4:99.Article Pubmed
[46]Kogoshi H, Tohda S, Fu L, Koyama T, Nara N. Effect of notch ligands on in vitro sensitivity to chemo-therapeutic drugs in leukemia and lymphoma cells. Oncol Rep. 2005; 14(3):723–726.Article Pubmed
[47]Damiano JS, Hazlehurst LA, Dalton WS. Cell adhesion-mediated drug resistance (CAM-DR) protects the K562 chronic myelogenous leukemia cell line from apoptosis induced by BCR/ABL inhibition, cytotoxic drugs, and gamma-irradiation. Leukemia. 2001; 15(8):1232–1239.Article Pubmed
[48]Kuželová K, Grebeňová D, Holoubek A, Röselová P, Obr A. Group I PAK inhibitor IPA-3 induces cell death and affects cell adhesivity to fibronectin in human hematopoietic cells. PLoS One. 2014; 9(3):e92560.Article Pubmed
[49]Aljitawi OS, Li D, Xiao Y, Zhang D, Ramachandran K, et al. A novel three-dimensional stromal-based model for in vitro chemotherapy sensitivity testing of leukemia cells. Leuk Lymphoma. 2014; 55(2):378–391.Article Pubmed
[50]Xishan Z, Ziying L, Jing D, Gang L. MicroRNA-320a acts as a tumor suppressor by targeting BCR/ABL oncogene in chronic myeloid leukemia. Sci Rep. 2015; 5:12460.Article Pubmed
[51]Chitteti BR, Bethel M, Voytik-Harbin SL, Kacena MA, Srour EF. In vitro construction of 2D and 3D simulations of the murine hematopoietic niche. Methods Mol Biol. 2013; 1035:43–56.Article Pubmed
 Copyright
© 2012-2019 NobleResearch Group. All Rights Reserved
Copyright
© 2012-2019 NobleResearch Group. All Rights Reserved